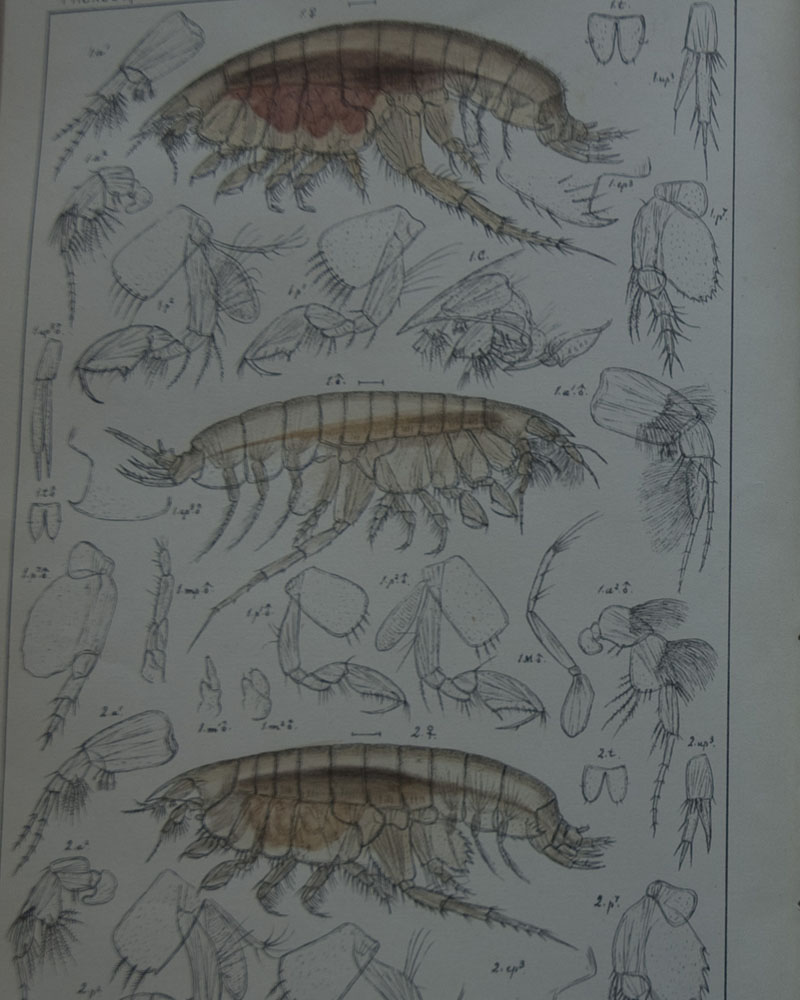
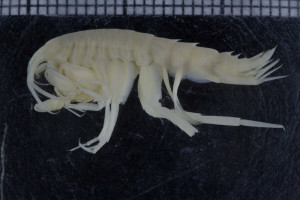
Den generelle amfipoden vi har blitt kjent med gjennom TangloppeTorsdag er for det meste avlang, flat sidelengs (som om den har blitt skvist mellom hendene dine når du gir applaus for hvor kule amfipoder er), og med bein som starter med en sideplate (coxalplate) som rett nok kan være stor eller liten, men som er en plate, og en bakkropp med tre par med halevedheng (uropoder) som hos de karakteristiske Gammarus artene.
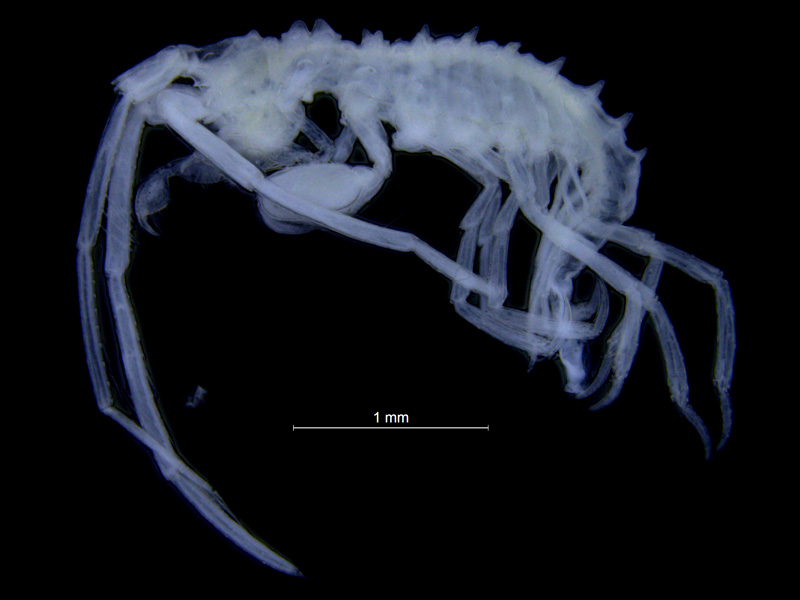
Laetmatophilus tuberculatus. Foto: AHS Tandberg
For noen grupper er det andre karakterer som gjør at vi kan si at det er amfipoder vi ser på. Laetmatophilus tuberculatus tilhører en av disse gruppene. (Det vil helt sikkert bli flere “uvanlige” amfipoder etterhvert, så stay tuned!) Det som gjør at vi allikevel kan kjenne dem igjen som amfipoder er det at de har frambeinpar som bøyes bakover og bakbeinpar som bøyes framover, i tillegg til at hele kroppen er leddet.
L. tuberculatus ser til forveksling ut som en litt lett piggete klump med lange tynne bein på, sideplatene på hvert bein er så små at de nesten ikke er til å se. På det nestfremste beinparet er det nestnederste leddet (som i gamle dager ble kalt “hånden”) utvidet, slik at det ser ut som boksehansker. Så mye at på en barneaktivitet med fargelegging av strektegninger av forskjellige arter jeg en gang hadde i en amfipodeutstilling, var det flere barn som farget dette leddet rødt – som røde boksehansker, og en jente ville kalle amfipoden sin for Rocky – etter den legendariske filmbokseren. Denne store hånden er mye mer utpreget hos hannene enn hos hunnene, men begge kjønn har slike boksehansker. Den karakteren som gjør at denne arten skiller seg mest ut fra den generelle amfipoden er allikevel det at de to bakerste kropssegmentene er sammenvokste, og de har bare to par halevedheng (uropoder), og det bakerste av de to den har er så redusert i form at det er lett å tro at det bare er et par uropoder hos denne slekten.

Laetmatophilus tuberculatus. Foto: K. Kongshavn
Dette er en art som lever på bløte mudderbunner, der er den observert mens den gjemmer seg under skjellrester og lignende skulesteder. Vi mener de spiser detritus (dødt, organisk “nedfall”), og med lange, hårete antenner kan vi også se for oss at en del av matinnsamlingen er basert på å vifte med antennene og så spise det som fester seg der. Den svenske zoologen Paul Enequist observerte i sitt arbeid en del nærståene arter i akvarium over lengre tid. Han beskrev hvordan de holdt seg fast med bakbeina mens de stilte framkroppen inn slik at vannstrømmen kom “rett i fleisen”, og så spredte de antennene ut så bredt de kunne, og lepjet i seg det som festet seg på dem. Enequists laboratorie-undersøkelser er fremdeles unike i amfipode-sammenheng: det er veldig få andre som har systematisk undersøkt levemåten til våre små venner. (I de seineste tiårene har det kommet noen få nye studier av levemåten et par arter – kanskje kommer det mer framover?)
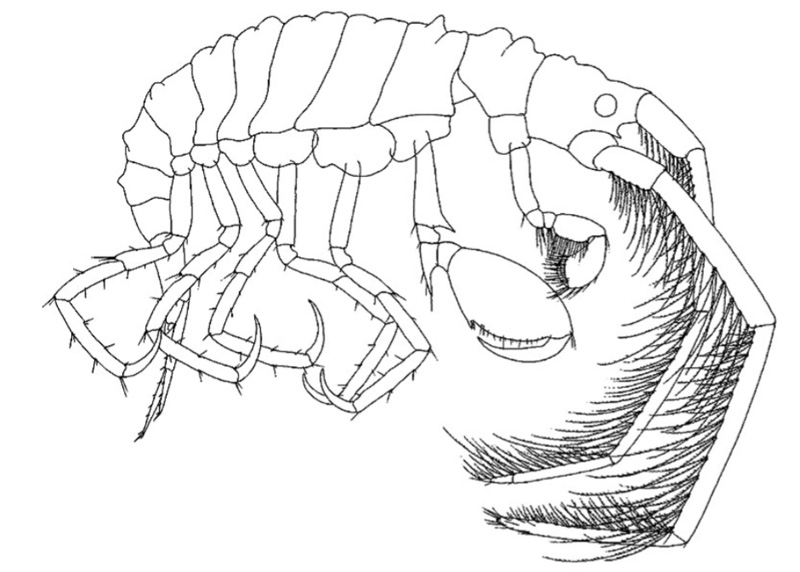
Laetmatophilus tuberculatus tegnet av Lincoln (1979).
Hvorfor ser Laetmatophilus tuberculatus så anderledes ut fra de fleste andre amfipodene? Det er flere grupper amfipoder som har slik redusert bakkropp (at få eller mange vedheng ikke finnes hos disse gruppene), og vi tror at dette kan ha skjedd tidlig i utviklingen av amfipodene. Et resultat av å ha en “enklere” bakkropp er at det blir lettere å holde seg fast i omkringliggende strukturer (tang, skjellrester, andre dyr) med bakbeina, og slik kan frambeina brukes til andre oppgaver som for eksempel matinnsamling eller kanskje til og med til å holde fast partnere i heftige stunder?
Det tidlige evolusjonspresset på amfipodene er ganske lite studert, og fremdeles diskuterer vi teorier uten helt å bli enige om hvilke vi synes har best støtte. Selv om vi bare er mellom 80 og 100 forskere på verdensbasis, har vi mange teorier vi undersøker, og flere forskjellige metoder vi tester teoriene med. Analyser av arvestoff (DNA) er selvsagt ønskelig, men det gjelder å få tak i nok representanter fra forskjellige arter til at det har noe for seg å sammenligne resultatene.

Laetmatophilus tuberculatus, dette er et av individene vi har DNA strekkode fra. Foto: K. Kongshavn
Gjennom NorAmph og NorBOL prosjektene her på Universitetsmuseet i Bergen har vi samlet inn strekkode-DNA for Laetmatophilus tuberculatus, som er den arten som finnes i våre farvann av 14 arter på verdensbasis for Laetmatophilus. De fleste av de resterende 13 andre kjente artene har ikke noe registrert DNA-materiale enda, for ikke å snakke om det vi med morfologiske (basert på utseende) metoder kan tenke oss at kanskje er nærtstående grupper. Når vi får nok slikt materiale kan vi sjekke om de forskjellige metodene våre gir resultater som ligner på hverandre. Slik kan vi bringe forståelsen av amfipodenes utvikling litt videre.
Så, det er bare å brette opp ermene og samle inn og analysere videre. Planen for min sommer er å samle inn materiale fra områder dypt i Norskehavet – kanskje ikke Laetmatophilus tuberculatus i denne omgang, de bor litt grunnere. Registrerte funn er på mellom 40 og 600m. Det er mange andre andre arter amfipoder å samle inn, så jeg er ikke redd for at det skal bli for lite å gjøre på tokt eller i laben til høsten.
Har du planer om å se på amfipoder i løpet av sommeren? Det er mange å velge mellom – både på stranden, i fjæra, fra ferskvann og fra langt nede i havet. Ta med deg bøtte og litt tålmodignet, kanskje blir det begynnelsen på slike studier Enequist holdt på med? Sommerutgavene av TangloppeTorsdag vil fortelle litt om noen av amfipodene du kan finne – følg med!
Anne Helene
Litteratur:
Enequist, P. 1949. Studies on the Soft-Bottom Amphipods of the Skagerak. Zoologiska Bidrag från Uppsala, 28.
Laubitz, D.R. 1983. A revision of the family Podoceridae (Amphipoda: Gammaridea). Australian Museum Memoir vol 18, 77-86.
Lincoln, R.J. 1979. British Marine Amphipoda: Gammaridea. British Museum, Natural History.